Outline of Cell cell communication.png - Wikimedia Commons
Homeostasis is the most important part of physiology, and all control systems try to keep it stable. The various control systems cannot function without the ability of cells to communicate with one another. This shows how the flow of information between cells, tissues, and organs is a basic principle of physiology that makes it possible for physiological processes to work together. Communication with the outside world is done through a number of pathways that take in and process signals from the outside world, from other cells in the same organism, and from different parts of the cell itself.
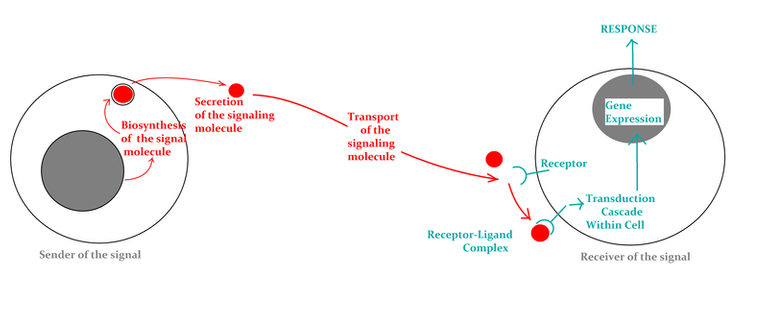
A signal is a piece of encoded information that starts or triggers a reaction. So, cell signaling is the process and method by which cells make and send encoded data to control and coordinate a number of bodily functions. Through the use of cell signaling, a single cell can specifically affect how other cells behave. It aids in the regulation of metabolic adjustments, cell reproduction, growth, differentiation, and development, among other things.
GENERATION OF SIGNALS
All physiological signs are produced as a consequence of alterations that may take place inside the cell, in the cell's internal surroundings, or in the outside world. Not all alterations to the physical or chemical environment are potent enough to cause bodily signs. Threshold refers to the moment at which a stimulus is just strong enough to be recognized by the cell. The visible spectrum is the range of wavelengths that activate photoreceptor cells and is, for example, undetectable by electromagnetic waves with wavelength lower than 380 nanometers or longer than 760 nanometers.
RECEPTORS
The target cell needs to have a sensor for the signal in order to recognize it. A receptor is a special molecule (protein or glycoprotein) or molecular compound that can recognize a certain signal and start a chain of events that leads to a certain reaction. The configuration of the receptor is believed to change in reaction to signal interaction, changing how the receptor molecule interacts with other molecules close by in the response pathway. On the other hand, a biomolecule or ion that precisely and preferentially attaches to a receptor and forms a complex with it in order to trigger a biological reaction is known as a ligand. A target cell is any cell that attaches a given ligand specifically and preferentially using receptors, whether or not a biochemical or physiological reaction has been found. It could be a hormone, an organic substance, or an inorganic substance.
Various receptor types
a. The plasma membrane receptors
Transmembrane proteins, which cover the full length of the membrane, are what make up plasma membrane receptors. Like other transmembrane proteins, plasma membrane receptors have hydrophobic segments that are located inside the membrane, hydrophilic segments that protrude into the external fluid from the membrane, and hydrophobic segments that protrude into the internal fluid. The extracellular sections of the receptor are involved in binding to incoming chemical signals (ligands); the intracellular regions of the receptor are engaged in signal transmission activities (to be discussed later). Examples include neurotransmitters and protein hormones like prolactin, growth hormone, insulin, and parathormone (acetylcholine, norepinephrine).
b. Intracellular receptors
Intracellular receptors, on the other hand, have a very distinct shape and are not found in membranes; instead, they are found in the cytosol or the cell nucleus. However, they share features with plasma membrane receptors, such as a section that attaches the messenger and additional segments that serve as regulatory sites. In contrast to plasma membrane receptors, they additionally have a section that attaches to DNA. One significant difference between the two main categories of receptors is that while all intracellular receptors transduce signals through interactions with genes, plasma membrane receptors can do so without doing so with DNA. Steroid hormones (cortisol, oestrogen, testosterone, aldosterone, etc.) and thyroxin are two examples.
Receptor-Ligand Relationships
a. Specificity: Even though a particular chemical messenger may come into contact with a variety of various cell kinds, it only affects some of them. This is a result of the various receptor kinds that exist in different tissues. Only a small number of cell types, or sometimes just one, make the receptor that is needed to attach a certain chemical message.
b. Affinity: The strength of a specific messenger's binding to its receptor depends on the receptor's affinity for that messenger. When it comes to attaching to a messenger, a receptor with a high affinity will need less of it than a receptor with a low affinity.
c. Saturation: When the amount of a messenger outside the cell goes up, more messenger molecules fill up the receptors. This makes the cell respond more to the messenger. However, there is a limit to this responsiveness because there are only a limited number of receptors, and they eventually reach complete saturation.
d. Competition: The capacity of a substance to outbid a naturally occurring ligand for attaching to a receptor is referred to as "competition." Competition underpins the action of many drugs and usually happens with messengers that are structurally comparable to one another.
CELL SIGNALING TYPES
There are five main kinds of cell signaling, depending on the location of the chemical message and the separation between the inducer and target cells.
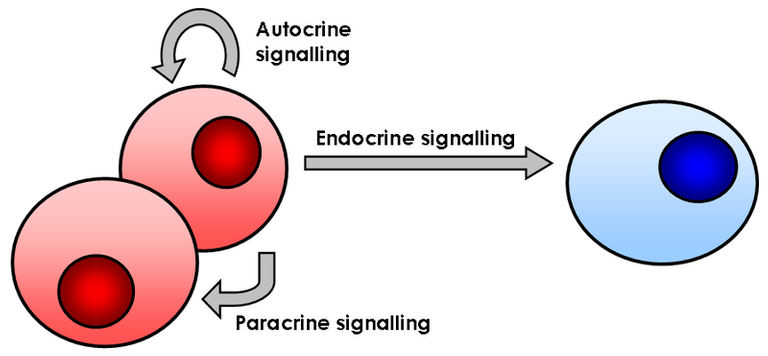
Juxtacrine Signaling: A juxtacrine signal is sent when proteins from the initiating cell bind to receptor proteins on nearby cells that can respond. The cell that produces the inducer does not let it spread. Juxtacrine reactions come in three different flavors. A protein on one type of cell attaches to its receptor on the other type of cell in the first type. In the second type, a binding reaction takes place when an extracellular matrix released by one cell binds to a receptor on another cell. The third type is when signals are sent directly from the cytoplasm of one cell to the cytoplasm of a nearby cell through tiny channels.
Paracrine Signaling: This type of cell communication happens when the cell that sends the signal is close to the cell it affects. Some molecules that send signals lose their effectiveness very quickly, so they can only be used in the immediate area. Others only affect nearby cells because they are quickly taken up, leaving few to move farther, or because the extracellular matrix keeps them from moving freely. Paracrine signaling components include growth hormones and clotting proteins.
Autocrine Signaling: The hormone interacts with the cell that makes it as well as cells that are similar to it after coming into contact with the interstitial fluid. Intracrine transmission occurs when there is no interaction with the interstitial fluid. For example, cytokines and growth factors help cells form and change into different types of cells.
Neurocrine Signaling: This happens when neurons generate chemicals that are delivered to their target cells via the axons (axoplasmic flow). Neuromuscular transmissions and axo-axonal transmissions are common instances.
Hormone Signaling: This is distant communication. Chemicals are discharged straight into extracellular spaces, where they are carried to distant locations by the blood (target tissues). Pituitary and adrenal chemicals are two examples.
File: Cell signalling.png - Wikimedia Commons
TRANSDUCTION OF SIGNALS
Transduction generally refers to the change of one type of energy into another. In terms of cell signaling, signal transduction is the process by which physiological information is turned into a certain cellular response. Signal transduction paths are the different sets of events that link the activation of a receptor to a reaction in a cell. The term "pathways" refers to the cell-specific processes connected to various messages. The trajectory or order of signal transduction paths can be thought of as a molecular circuit and is generally comparable. The following is the sequence:
Membrane sensors transmit information that comes from the outside to the body of the cell.
An inherent membrane protein with extracellular and intracellular regions makes up such a receptor. The signal molecule, also called the ligand, is found in the binding site of the extracellular domain. The contact between the ligand and the receptor changes the tertiary or quaternary structure of the receptor, including the area inside the cell. Because they only affect a small number of receptor molecules in the cell membrane, these changes in structure are not enough to cause the right reaction. Before the cell's metabolism can be changed, the information carried by the ligand, which is also called the main messenger, must be changed into another form.
The receptor-ligand combination transmits information through second messengers.
The next stage in the molecular communication pathway involves changes in the concentration of small molecules known as "second messengers." Cyclic AMP and cyclic GMP, calcium ions, inositol 1,4,5-triphosphate (IP3), and diacylglycerol are second messengers that are particularly significant (DAG). Using second-class couriers has a number of repercussions. First, second messengers often have free access to other parts of the cell, like the nucleus, where they can change how genes are translated and other things. Second, the production of second messages has the potential to greatly amplify the signal. Most of the time, enzymes or membrane channels have to be turned on for second messengers to be made. Each macromolecule that is turned on can cause many second messengers to be made inside the cell. So, even a small amount of information in the environment, like one molecule, can cause a big signal and response inside the cell.
A frequent method of information transmission is protein phosphorylation.
Protein kinases are frequently activated by second messengers to trigger reactions. Changes in the amount of unbound second messengers are converted into changes in covalent protein structures by protein kinases.
The signal is cut off
One method for stopping a signaling process is protein phosphatase. Once the signaling processes have begun and the information has been sent to change other cellular processes, they must be stopped.
References
- Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K., & Walter, P. (2002, January 1). General Principles of Cell Communication - Molecular Biology of the Cell - NCBI Bookshelf. General Principles of Cell Communication - Molecular Biology of the Cell - NCBI Bookshelf. https://www.ncbi.nlm.nih.gov/books/NBK26813/
- Cell Signaling | Learn Science at Scitable. (n.d.). Cell Signaling | Learn Science at Scitable. https://www.nature.com/scitable/topicpage/cell-signaling-14047077/
- Cell Communication | Learn Science at Scitable. (n.d.). Cell Communication | Learn Science at Scitable. https://www.nature.com/scitable/topic/cell-communication-14122659/
- Mattaini, K. (n.d.). Chapter 9. Cell Communication – Introduction to Molecular and Cell Biology. Chapter 9. Cell Communication – Introduction to Molecular and Cell Biology. https://rwu.pressbooks.pub/bio103/chapter/cell-communication/
- Introduction to cell signaling (article) | Khan Academy. (n.d.). Khan Academy. https://www.khanacademy.org/science/ap-biology/cell-communication-and-cell-cycle/cell-communication/a/introduction-to-cell-signaling
- Cell Communication and Signaling. (n.d.). BioMed Central. https://biosignaling.biomedcentral.com/
Absolute great post
Thanks so much for stopping by
Your article is good!
By the way, I have a hard time understanding your professional knowledge!!😅Dear @jsalvage !
Smile,, I took time to simplify the article though
Thanks for your contribution to the STEMsocial community. Feel free to join us on discord to get to know the rest of us!
Please consider delegating to the @stemsocial account (85% of the curation rewards are returned).
Thanks for including @stemsocial as a beneficiary, which gives you stronger support.
This is a great article
So much knowledge to have here
I'm glad you liked it .